



业务咨询

![]() 发布时间:2023-07-21
发布时间:2023-07-21
![]() 环特生物
环特生物
![]() 浏览次数:6539
浏览次数:6539

编者按
不同类器官需要不同的生长环境,尤其是培养基中的生长因子,需要精心调配,确定使用的生长因子种类与浓度。那么,哪些类器官需要哪些生长因子呢?该如何确定使用哪些生长因子呢?
今天,我们特别回顾一项于2022年发表在《Cell Stem Cell》的研究——《Optimized human intestinal organoid model reveals interleukin-22-dependency of paneth cell formation》,该研究通过调查白细胞介素(IL)-22这一因子对类器官生长的作用,优化了人类小肠类器官(hSIO)的培养方法,揭示了IL-22不直接控制肠道干细胞的再生能力,而是会增加潘氏细胞数量以及促进肠道抗菌肽(AMP)的表达。
一、 研究背景
白细胞介素(IL)-22由免疫细胞产生,主要作用于上皮细胞,可视为免疫系统和表达IL-22共受体复合物的特殊组织细胞之间的主要交流渠道。IL-22与肠道生理的多个方面有关,它被认为是粘膜愈合的重要驱动力。一项关键研究(主要基于小鼠类器官实验)提出,IL-22通过直接增加肠道干细胞(ISCs)的数量和增殖活性来促进组织再生[1]。
然而,最近的小鼠研究表明,IL-22 通过诱导过渡增殖细胞 (TA) 的增殖来促进再生,同时抑制 ISC 扩增甚至诱导 ISC 细胞凋亡[2]。IL-22已被广泛评估为炎症性肠病(IBD)的潜在治疗方法[3]。因为据报道,IL-22 可通过促进杯状细胞分化和诱导粘蛋白分泌,增强肠道屏障对病原体防护[4]、促进抗菌蛋白(AMP)[5]和补体因子[6]的产生。
IL-22在IBD中的作用仍未完全阐明。在临床前小鼠模型中,阻断IL-22的产生或与受体的结合可改善肠道炎症[7]。与之相反的观点是,IL-22或IL-22受体的缺乏导致结肠炎恶化,而用IL-22-Fc融合蛋白治疗改善了野生型小鼠的炎症[8]。此外,在IBD患者中,观察到屏障功能障碍和疾病进展的持续症状与IBD患者的IL-22水平升高有关[9],表明IL-22在IBD中的致病作用。然而,另一项研究揭示IL-22在IBD患者中具有保护作用[10]。
小肠类器官(SIO)提供了一种体外模型,用于研究在没有其他因素的情况下生物调节剂对原代上皮细胞的直接影响[11]。虽然人小肠类器官(hSIO)的培养条件已有报道[12],然而人类肠道组织的细胞类型多样性和三维 (3D) 结构尚未完全概括。值得注意的是,潘氏细胞可产生肠道AMPs(如溶菌酶和防御素),这在调节宿主防御方面至关重要[13]。但人肠道中潘氏细胞的分化过程尚未确定。因此本文中,研究者建立了一个优化的hSIO培养方案,探测了IL-22对人小肠上皮中各种上皮细胞类型的诱导作用以及基因表达的直接影响。
二、 研究结果
1、优化的人小肠类器官培养方法本研究
基于经典研究中的传统培养方法[14],开发了一套“两步法”hSIO培养方案,与传统扩增培养基相比,本研究优化后的第一步培养基(称为“模式化培养基”)中去除了抑制剂SB202190、前列腺素E2和烟酰胺,又添加了抑制剂CHIR99021和IL-22;第二步培养基(称为“成熟化培养基”)在第一步的基础上降低了外源WNT信号。本研究的模式化培养基驱动hSIO向分泌谱系分化。
模式化14天后,将模式化培养基替换为成熟化培养基,以支持具有广泛隐窝状出芽结构的类器官扩增。这样培养得到的hSIO中,潘氏细胞、杯状细胞和肠内分泌细胞分散于整个类器官结构中。用免疫组织化学标志物染色和透射电子显微镜的形态学表征证实了这些分泌细胞类型的成熟。
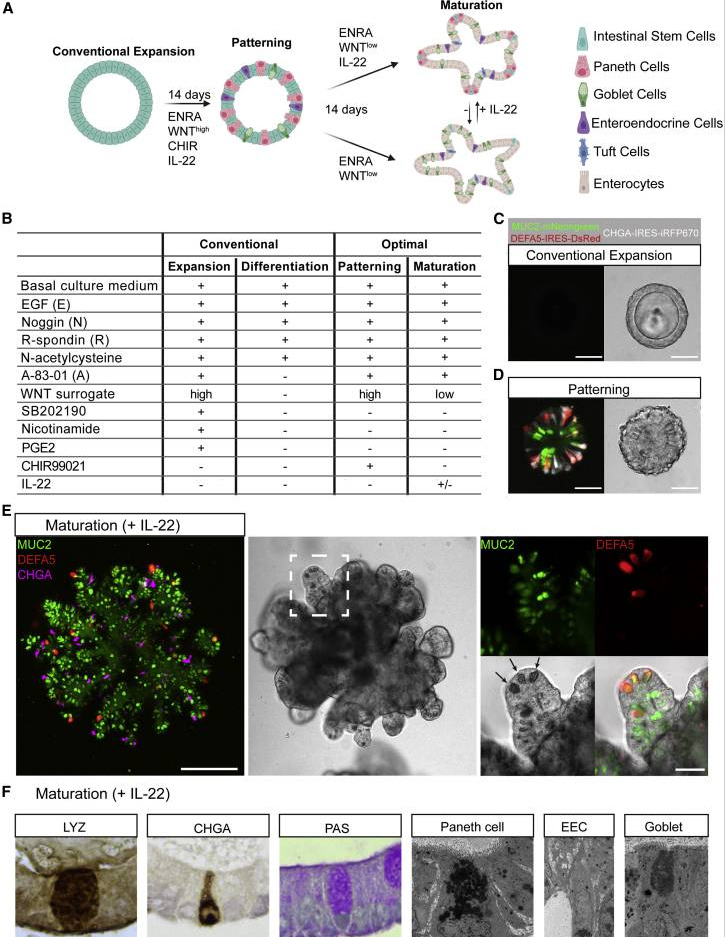
原文图1. 具有多分化能力的hSIO的培养
为了对这种hSIO的细胞多样性进行充分了解,研究人员对成熟的hSIO(在IL-22存在下生长了14天)进行了单细胞RNA测序(scRNA-seq)分析。本研究的数据捕获了1283个细胞,这些细胞聚集成10个群体,包括ISC、TA的两个亚簇、早期和晚期肠细胞(enterocytes,肠道吸收型上皮细胞)、分泌型祖细胞、杯状细胞、潘氏细胞、肠内分泌细胞(EEC)和簇细胞。
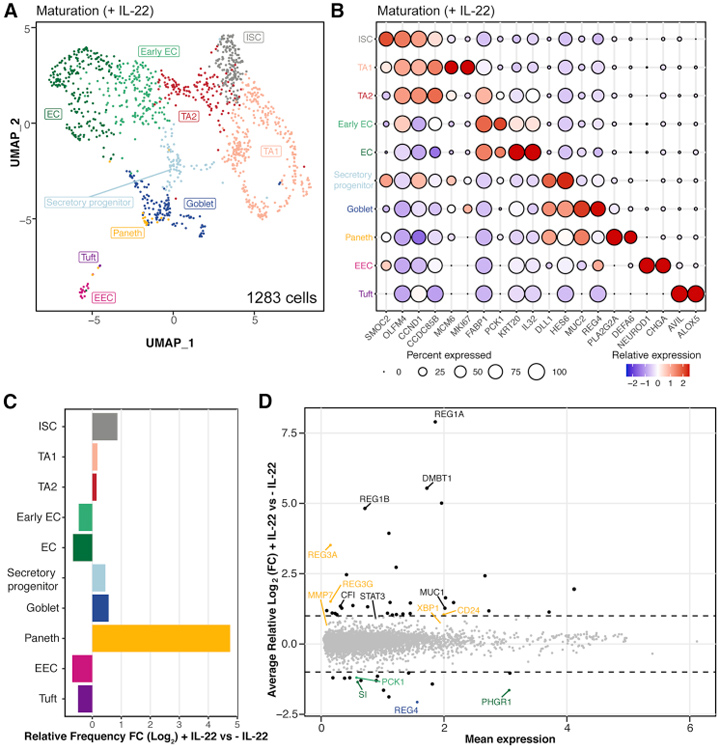
图2. IL-22调控hSIO的细胞多样性
2、IL-22并不会促进肠道再生
在培养基中添加IL-22,并不会影响ISC的绝对数量,虽然ISC的相对比例增加了,但却减少了活细胞的总数,同时显著降低了类器官形成的效率。此外,在培养基中添加IL-22减少了类器官出芽并增加了细胞死亡,表明IL-22在hSIO的长期培养中起到了有害作用。
总的来说,IL-22既不增强ISC的扩增,也不增强hSIO的生长。scRNA-seq结果表明,IL-22降低了(早期)enterocytes生长比例。为了证实这一观察结果,研究人员检测了ANPEP和CYP3A4基因(仅表达于enterocytes),结果显示,IL-22确实抑制了enterocytes相关基因如ANPEP和CYP3A4。
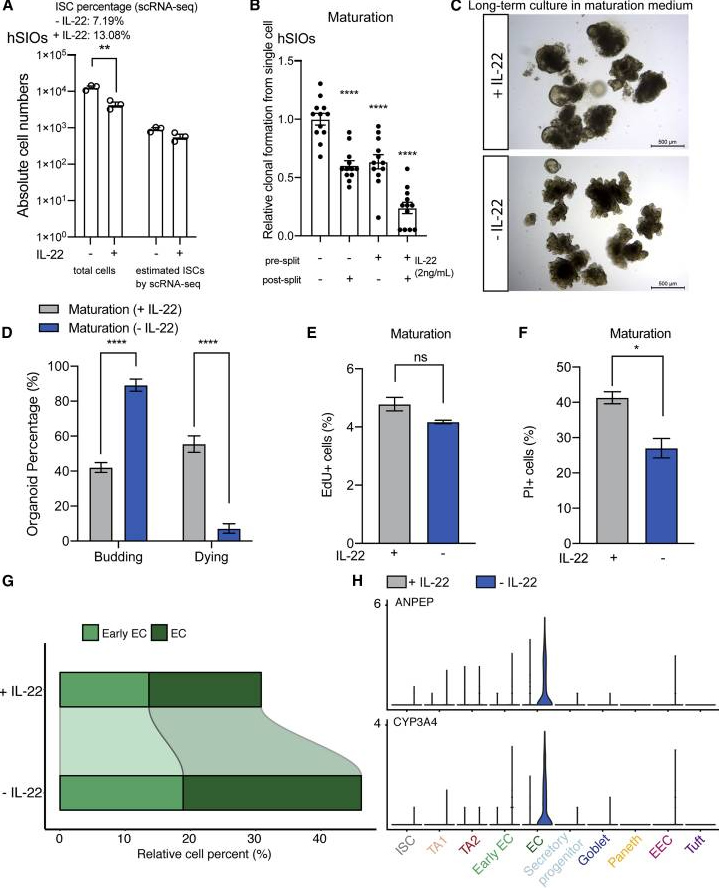
图3. IL-22抑制hSIO的生长
3、IL-22诱导人潘氏细胞分化
用去除IL-22的成熟化培养基培养获得的hSIO中几乎不存在潘氏细胞,表明IL-22起到促进潘氏细胞分化的作用。而用含有IL-22的成熟化培养基培养hSIO,潘氏细胞在一周内出现,而再次去除IL-22会导致潘氏细胞在两周内消失,符合潘氏细胞的寿命周期。
类器官中杯状细胞和EEC的百分比不受IL-22的影响,表明IL-22特异性诱导潘氏细胞分化,而不是驱动整体分泌谱系。hSIO可以每周传代一次,并在没有IL-22的成熟化培养基中扩增>12个月,之后潘氏细胞仍然可以被IL-22有效地诱导。这种可逆表现将IL-22信号传导确定为潘氏细胞稳态的关键介质。
为了探测潘氏细胞的起源,本研究进行了基于图像的实时谱系跟踪,发现新诱导的潘氏细胞均来源于MUC2+祖细胞,支持潘氏细胞和杯状细胞共享共同分泌祖细胞群体的观点。
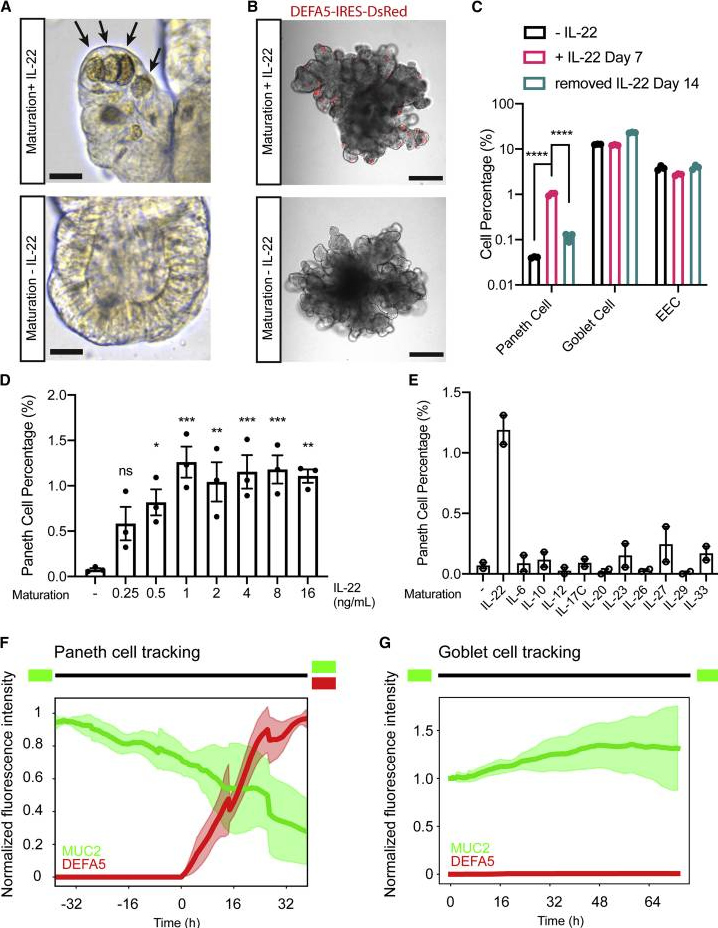
图4. IL-22诱导人潘氏细胞分化
4、IL10RB敲除导致hSIO中的潘氏细胞缺乏
IL-22受体基因 IL10RB的突变导致罕见的遗传性 IBD。本研究创建了两个IL10RB功能丧失的突变基因W159∗和 W204∗。本研究表明,携带这两种突变基因中的任何一种的hSIO,即使用含有IL-22的成熟化培养基培养后,也是缺乏潘氏细胞的,而杯状细胞和EEC不受影响。
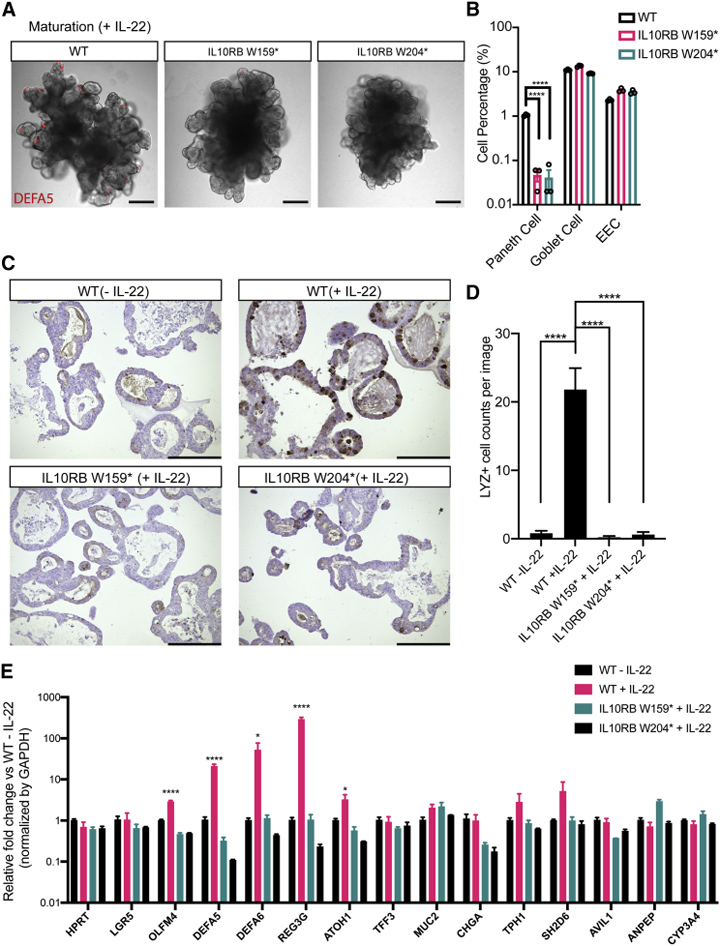
图5. IL10RB是IL-22诱导的潘氏细胞分化所必需的
5、PI3K/AKT/mTOR通路介导IL-22下游的人潘氏细胞分化
为了了解 IL-22诱导的潘氏细胞分化的机制,研究人员通过小分子抑制剂检测了 IL-22下游信号传导途径 STAT3和 PI3K/AKT/ mTOR。与未处理的类器官相比,在用含有IL-22成熟化培养基培养的hSIO中,通过BP-1-102阻断STAT3活化对 DEFA5阳性潘氏细胞的比例没有影响。
相反,PI3K抑制剂LY294002或 AKT抑制剂MK-2206显著降低了潘氏细胞的存活率。更值得注意的是,抑制mTOR几乎完全阻断了IL-22诱导的潘氏细胞分化。PI3K,AKT 或 mTOR 抑制剂以剂量依赖性方式降低了潘氏细胞存活率,而杯状细胞存活率没有显著改变。这些结果表明 PI3K/AKT/mTOR 信号通路特异性地驱动潘氏细胞分化而不影响其他分泌谱系。
本研究进一步在四种不同条件下对成熟培养基培养的类器官进行定量蛋白质组分析:-IL-22,+IL-22,+IL-22与STAT3抑制剂BP-1-102共用,+IL-22与mTOR 抑制剂雷帕霉素共用。IL-22显著上调了几种潘氏细胞蛋白,包括DEFA5,DEFA6,Reg3A和ITLN2,但在抑制mTOR时,这些蛋白质的增加显著被挽救。这些蛋白质组学数据揭示了mTOR介导IL-22促进潘氏细胞分化的进一步证据。
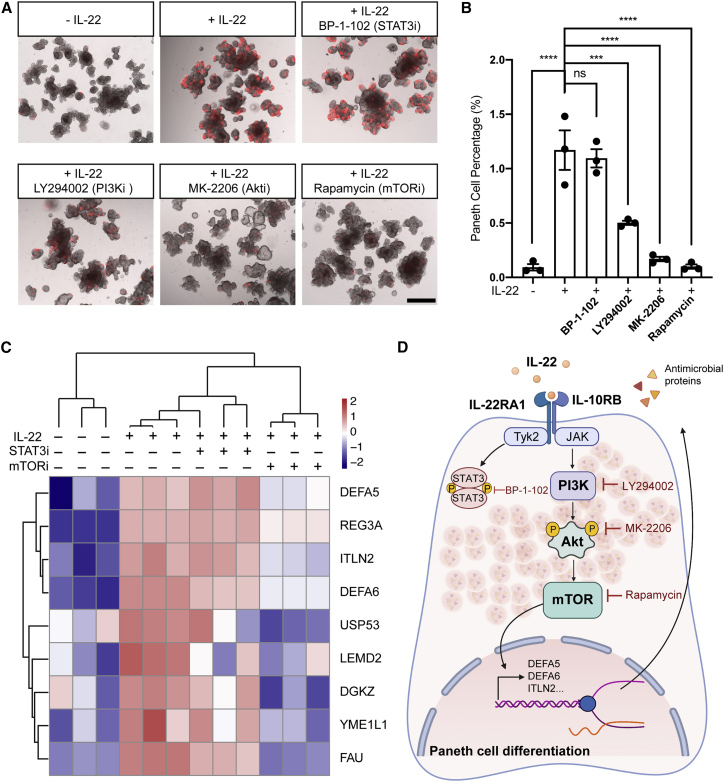
图6. PI3K-mTOR信号传导介导IL-22下游的潘氏细胞分化
6、IL-22诱导的AMP表达不仅限于潘氏细胞
除了一些潘氏细胞特异性基因(例如DEFA6和ITLN2)外,前50个IL-22上调的基因在所有细胞类型中均有表达。这些基因包括AMP(Reg1A、Reg1B和DMBT1)、补体因子(如CFI)和粘蛋白(如MUC1)。
事实上,IL-22共同受体IL10RB 和 IL22RA 在 hSIO 中由所有上皮细胞类型表达。在24小时内通过 IL-22刺激显著诱导这些基因(如Reg1A、Reg1B 和 DMBT1)的表达。这些AMP基因的表达不是由与潘氏细胞分化相同的信号传导途径驱动的,因为与潘氏细胞标记不同,这些基因的表达不受雷帕霉素处理的影响。在人小肠切片上Reg1A/B和DMBT1的组织学分析显示,它们在隐窝的下半部分表达最高,并不仅限于潘氏细胞。
总的来说,这些数据表明IL-22可能作为隐窝底部宿主防御的生态位因子,其中 IL-22不仅诱导潘氏细胞分化,而且诱导肠干细胞及其子代中AMPs 的表达。
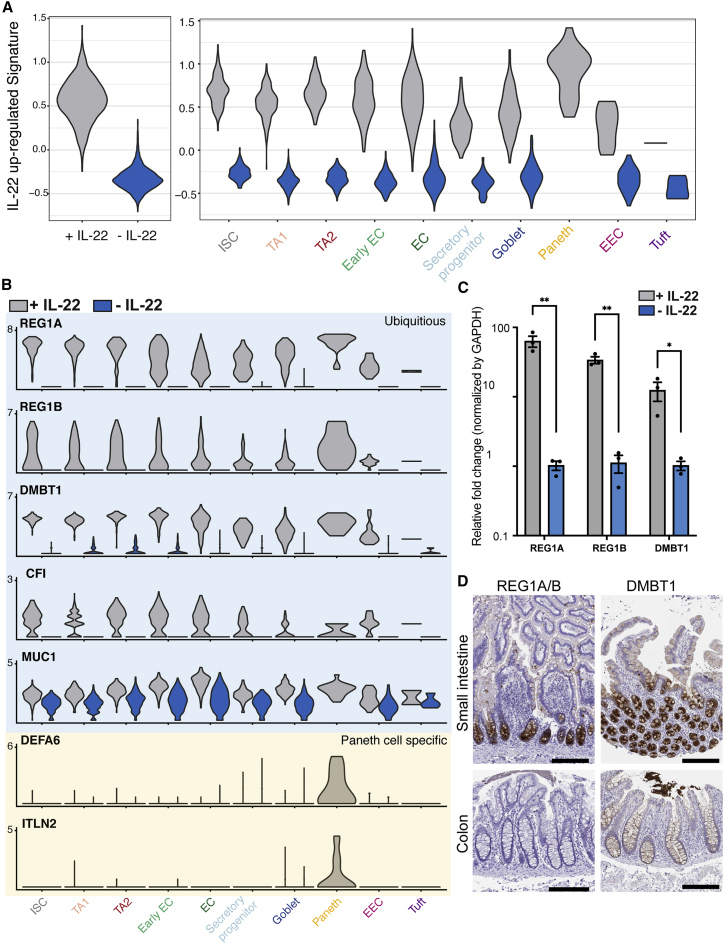
图7. IL-22诱导宿主防御基因表达
三、 编者点评
本研究描述了hSIO优化培养系统的建立,并且通过探究IL-22对诱导人小肠上皮的各种上皮细胞的直接影响,表明优化的培养条件显著改善了人肠上皮的离体建模,并显著拓宽了人肠道类器官的应用;确定了IL-22在通过PI3K/AKT/mTOR通路诱导人潘氏细胞分化中起到重要作用。本研究将IL-22确定为在高Wnt信号环境中体外优化hSIO中潘氏细胞形成的诱导剂,但不能证明IL-22是体内潘氏细胞形成的唯一必需诱导剂。
作为斑马鱼生物技术的全球领导者,环特生物搭建了“斑马鱼、类器官、哺乳动物、人体”四位一体的综合技术服务体系,开展科研及研究服务、智慧实验室建设和精准医疗三大业务。目前,环特类器官平台已成功搭建多种肿瘤及正常类器官的培养平台,已有多种肿瘤类器官培养试剂盒和试剂在售,欢迎各位读者垂询!
参考文献
[1] Lindemans C.A., Calafiore M., Mertelsmann A.M., O'Connor M.H., Dudakov J.A., Jenq R.R., Velardi E., Young L.F., Smith O.M., Lawrence G., et al. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial regeneration. Nature. 2015;528:560–564.
[2] Zha J.M., Li H.S., Lin Q., Kuo W.T., Jiang Z.H., Tsai P.Y., Ding N., Wu J., Xu S.F., Wang Y.T., et al. Interleukin 22 Expands Transit-Amplifying Cells While Depleting Lgr5(+) Stem cells via inhibition of Wnt and notch signaling. Cell. Mol. Gastroenterol. Hepatol. 2019;7:255–274.
[3] Rothenberg M.E., Wang Y., Lekkerkerker A., Danilenko D.M., Maciuca R., Erickson R., Herman A., Stefanich E., Lu T.T. Randomized phase I healthy volunteer study of UTTR1147A (IL-22Fc): a potential therapy for epithelial injury. Clin. Pharmacol. Ther. 2019;105:177–189.
[4] Layunta E., Jäverfelt S., Dolan B., Arike L., Pelaseyed T. IL-22 promotes the formation of a MUC17 glycocalyx barrier in the postnatal small intestine during weaning. Cell Rep. 2021;34:108757.
[5] Gaudino S.J., Beaupre M., Lin X., Joshi P., Rathi S., McLaughlin P.A., Kempen C., Mehta N., Eskiocak O., Yueh B., et al. IL-22 receptor signaling in Paneth cells is critical for their maturation, microbiota colonization, Th17-related immune responses, and anti-Salmonella immunity. Mucosal Immunol. 2021;14:389–401.
[6] Hasegawa M., Yada S., Liu M.Z., Kamada N., Muñoz-Planillo R., Do N., Núñez G., Inohara N. Interleukin-22 regulates the complement system to promote resistance against pathobionts after pathogen-induced intestinal damage. Immunity. 2014;41:620–632.
[7] Eken A., Singh A.K., Treuting P.M., Oukka M. IL-23R+ innate lymphoid cells induce colitis via interleukin-22-dependent mechanism. Mucosal Immunol. 2014;7:143–154.
[8] Cox J.H., Kljavin N.M., Ota N., Leonard J., Roose-Girma M., Diehl L., Ouyang W., Ghilardi N. Opposing consequences of IL-23 signaling mediated by innate and adaptive cells in chemically induced colitis in mice. Mucosal Immunol. 2012;5:99–109.
[9] Schmechel S., Konrad A., Diegelmann J., Glas J., Wetzke M., Paschos E., Lohse P., Göke B., Brand S. Linking genetic susceptibility to Crohn's disease with Th17 cell function: IL-22 serum levels are increased in Crohn's disease and correlate with disease activity and IL23R genotype status. Inflamm. Bowel Dis. 2008;14:204–212.
[10] Pelczar P., Witkowski M., Perez L.G., Kempski J., Hammel A.G., Brockmann L., Kleinschmidt D., Wende S., Haueis C., Bedke T., et al. A pathogenic role for T cell-derived IL-22BP in inflammatory bowel disease. Science. 2016;354:358–362.
[11] Sato T., Vries R.G., Snippert H.J., van de Wetering M., Barker N., Stange D.E., van Es J.H., Abo A., Kujala P., Peters P.J., Clevers H. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 2009;459:262–265.
[12] Fujii M., Matano M., Toshimitsu K., Takano A., Mikami Y., Nishikori S., Sugimoto S., Sato T. Human intestinal organoids maintain self-renewal capacity and cellular diversity in niche-inspired culture condition. Cell Stem Cell. 2018;23:787–793.e6.
[13] Bevins C.L., Salzman N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011;9:356–368.
[14] Sato, T., Stange, D.E., Ferrante, M., et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology, 2011, 141,1762–1772.
